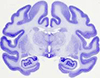
Imbedded in the structure of tiny gyrus, indusium griseum, they are, in fact, supracallosal fibers of the fornix, previously called fornix longus. 

In in vivo experiments, Abeta(25-35) was administered into the indusium griseum or the third ventricle and several parameters were measured 7 days later to evaluate potential Abeta(25-35)/BDNF interactions, i.e.
High-density specific binding was found in discrete areas of the brain and spinal cord, namely in the limbic system (hippocampal formation, septohippocampal nucleus, indusium griseum, hypothalamus, amygdaloid nuclei), superior colliculus, dorsal raphé, and substantia gelatinosa of the spinal cord.
Finally, a large population of somatostatin-negative GABAergic cells in stratum radiatum project to the molecular layers of the subiculum, presubiculum, retrosplenial cortex, and indusium griseum and fire rhythmically at high rates during theta oscillations but do not increase their firing during ripples.
The striae longitudinales mediales corporis callosi, or medial longitudinal striae of the corpus callosum, are two slender bands of myelinated fibers that form longitudinal ridges in the indusium griseum on the superior aspect of each half of the corpus callosum.
We show that radial glia in dorsomedial cortex retract their apical endfeet at midneurogenesis and translocate to the overlaying pia, forming the indusium griseum.
In the present study, we describe the distribution of basic fibroblast growth factor-2 at postnatal days 0, 2, 6, 10, 12, 14, 18, 21 and 30 in the indusium griseum, the external capsule, the hippocampus, the medial prefrontal cortex, the motor cortex, the rostral migratory stream, and the subventricular zone.
Administration of biotinylated dextran amine into the vitreous body resulted in nerve cell body labeling in several structures: the supraoptic and paraventricular nuclei, the hippocampus (CA1, CA3), the dentate gyrus, the indusium griseum, the olfactory tubercle, and the medial habenula, all of them belong to the limbic system.
Immunohistochemical analysis of the rat brain shows a prominent localization of palmdelphin in the cerebral cortex, hippocampus, amygdala, septum, indusium griseum, piriform cortex, nucleus supraopticus, and nucleus of the lateral olfactory tract.
These defects include the loss of the septum and three specialized glial cell types, the indusium griseum glia, midline zipper glia, and glial wedge.
As in rodents, three cellular clusters surrounding the corpus callosum were identified by their expression of GFAP and VN (but not MAP-2 or NeuN) from 13 to at least 18 weeks postovulation (wpo): the glial wedge, the glia of the indusium griseum, and the midline sling.
The indusium griseum and its precallosal extension are usually considered poorly differentiated portions of the hippocampus.
Initially, many neurons are found throughout the CC, while at later ages they become restricted to the boundaries of the CC, and in the adult to the rostrum of the CC close to the septum pellucidum or to the indusium griseum.
Agenesis or apparent hypoplasia of the indusium griseum of the limbic-cingular system as an accompanying finding in the present cases may support this hypothesis.
The most intense signals were observed in the ependymal cell layer along the wall of the third ventricle in the hypothalamus, CA1 region of the hippocampus, indusium griseum and septohippocampal nucleus.
Three midline glial populations are found at the corticoseptal boundary: the glial wedge (GW), glia within the indusium griseum (IGG), and the midline zipper glia (MG).
In adult male rats, 5-HT(7) receptor immunoreactivity was detected in the septum, striatum, indusium griseum, tenia tecta, thalamus, hippocampus and hypothalamus in the forebrain as well as the pons and cerebellum.
Three midline structures, the glial wedge, glia within the indusium griseum, and the glial sling are involved in development of the corpus callosum (Silver et al., 1982; Silver and Ogawa, 1983; Shu and Richards, 2001). Glia within the indusium griseum and the glial wedge are greatly reduced or absent and consequently Slit2 expression is also reduced.
Further parts of the brain we analyzed included the mammillary bodies, the indusium griseum and the cingulate cortex.
The results showed that a single injection of 40 mg/kg METH caused neuronal death in several brain areas including the striatum, cortex (frontal, parietal, and piriform), indusium griseum, medial habenular nucleus, and hippocampus.
The first differentiates directly below the corpus callosum forming a wedge shaped structure (the glial wedge) and the second differentiates directly above the corpus callosum within the indusium griseum.
Furthermore, the indusium griseum and taenia tecta were always present and the cerebral cortical layers were well differentiated in Emx1 mutant mice.
However, CD had no effect on apoptosis in the indusium griseum (IG), a region of cortex dorsal to the septum.
We also observed strong staining of ORX-ir varicose terminals in areas outside the hypothalamus, including the cerebral cortex, medial groups of the thalamus, circumventricular organs (subfornical organ and area postrema), limbic system (hippocampus, amygdala, and indusium griseum), and brain stem (locus coeruleus and raphe nuclei).
The human indusium griseum (IG), the paired dorsal continuation of the hippocampus, was investigated with high-field (3.0T) MR imaging.
In the telencephalon, transient 5-HT immunolabeling is observed in neurons in the CA1 and CA3 fields of the hippocampus, the central amygdala, the indusium griseum, and the deep layers of the anterior cingulate and retrosplenial cortices.
Two techniques (Fluoro-Jade labeling and argyrophylia) showed that neuronal degeneration occurred in the indusium griseum, tenia tecta and fasciola cinerea within 5 days post-METH exposure in 70% of the mice.
The bFGF immunoreactivity (IR) was detected within the cytoplasm of neurons, except within the pyramidal neurons of hippocampal CA2 region, the fasciola cinerea and the indusium griseum, where bFGF IR was present in the nucleus.
The result showed that the following ipsilateral structures are sending fibers to these three portions: the prefrontal cortex, frontoparietal motor cortex, indusium griseum, dorsal endopiriform nucleus, lateral part of medial mammillary nucleus, nuclei of diagonal band of Broca, anterior pretectum, anterior part of caudate-putamen, hippocampal formation, anteroventral, anteromedial, lateroposterior, ventroposterior and dorsomedial thalamic nuclei.
The anatomic locations of these neurons were the hippocampus, cerebellar cortex, superior colliculus, indusium griseum, mammillary nucleus, cerebral cortex, and the dorsal laminae of the dorsal horns of the spinal cord.
A moderately high concentration of binding sites was observed in the hippocampal formation, especially in the molecular layer of Ammon's horn, in the superficial layers of the entorhinal cortex and in the indusium griseum.
The supracallosal gyrus (indusium griseum), hidden from view by the corpus callosum, and the paraterminal gyrus are remnants of the previously larger hippocampal formation.
In situ hybridization revealed that rIRK3 mRNA is distributed throughout the brain and occurs predominantly in the piriform cortex, indusium griseum, supraoptic nucleus, facial nucleus and cerebellar Purkinje cells..
In situ hybridization analysis of adult mouse brain sections indicates that bsk is expressed at high levels in the hippocampus, tenia tecta, indusium griseum, and the piriform cortex, major components of the limbic system that are important for learning and memory.
The three mRNA variants were differentially distributed within the hippocampal subfields, with the alpha form being highly enriched in CA2, dentate, the fasciculum cinereum, and the indusium griseum, whereas beta and gamma forms were evenly distributed through CA1-4.
The immunocytochemical features of the indusium griseum (IG) were compared with the corresponding hippocampus in 5 patients with Alzheimer's disease (AD) and 5 age-matched nondemented individuals using antibodies against beta-amyloid, the A68 protein (Alz-50 antibody), tau, ubiquitin and synapsin I.
The pars tuberalis of the pituitary gland, suprachiasmatic nuclei, ventromedial hypothalamic nuclei, tapetum, hippocampus, indusium griseum, cingulate gyrus, cortex and the choroid plexus were intensely labelled.
Likewise, regions containing 5-HT axons of passage (e.g., indusium griseum and lateral hypothalamus) appeared to be insensitive to the neurotoxic effects of MDMA on 5-HT neurons.
At postnatal day (pnd) 2, a high density of labelled MR mRNA was found in all pyramidal (CA1-4) and granular (dentate gyrus) cell fields of the hippocampal structure, the anterior hippocampus and indusium griseum, and cortex layer II.
The histopathology of the indusium griseum (IG), a displaced hippocampal anlage, was studied in five patients with Alzheimer's disease (AD) and five controls.
The highest density of immunoreactivity was seen in the olfactory bulb, septohippocampal nucleus, indusium griseum, islands of Calleja, intermediate part of the lateral septal nucleus, and Ammon's horn.
Other extra-hypothalamic regions displaying a low-to-moderate density of immunoreactive fibers were the indusium griseum, the hippocampus, the fimbria of the fornix, the subcommissural organ, the medial habenula and, occasionally, the medial periaqueductal gray.
Neurons in areas CA2, CA3, CA4, and in the dorsomedial subiculum and indusium griseum showed little or no nuclear labeling after 3H dexamethasone. With 3H corticosterone, strongest nuclear labeling was observed in neurons in area CA2 and in the dorsomedial subiculum and indusium griseum, followed by area CA1, then CA3 and CA4; the dentate gyrus contained scattered strongly labeled cells among cells with intermediate nuclear labeling.
Only a few loci of basic FGF synthesis are detected by in situ hybridization and include layers 2 and 6 of the medial (cingulate) cortex, the indusium griseum, fasciola cinereum, and field CA2 of the hippocampus.
A scattering of LHRH-IR fibers were also observed in several extrahypothalamic regions, notably the subfornical organ, indusium griseum, habenular complex, septohypothalamic nucleus, and amygdala..
The indusium griseum, which demonstrated the highest binding for corticosterone of all brain regions in the autoradiograms, was taken as reference and defined as 100% relative labeling (RL).
The rudimentary parts of the hippocampal formation, taenia tecta and indusium griseum, were found to have reciprocal ipsilateral connections with the dorsal portions of the lateral septal nucleus..
A quantitative histological examination of brains from mice aged 25, 28 and 31 months of age showed that there was no loss of neurons from the indusium griseum and the number of glia per 100 neurons also remained constant. Unlike the indusium griseum, in which there was a significant variation in the mitotic and pyknotic indices with age, the neostriatum showed no variation in the mitotic and pyknotic indices between 6 and 31 months.
Neurons in cat medial prefrontal cortex, anterior limbic cortex and possibly the indusium griseum were identified by antidromic invasion as having axonal projections towards the medial preoptic region, using both macro- and microstimulation techniques.
In addition, some immunoreactive axons were observed in the anterior hippocampal continuation and the indusium griseum.
The indusium griseum (IG) is an enigmatic cortical field classically felt to be a part of the hippocampus (HC).
It was postulated that the wave of neuron birth and release which gave rise to the isocortical plate was propagated beyond the isocortical boundary into mesocortical territory as far as the boundaries of the subiculum, indusium griseum and anterior hippocampal rudiment..
However, in contrast to pretreatment with aldosterone, pretreatment with corticosterone and 18-OH-DOC did not completely prevent the concentration of radioactivity in the cell nuclei of the indusium griseum. A similar profile of silver grain distribution to that noted with aldosterone was found for corticosterone except that with tritiated corticosterone the most intense concentration of radioactivity occurred in hippocampal areas CA1 and CA2 and not in the indusium griseum.
No staining was found in the gyrus fasciolaris, indusium griseum, or anterior hippocampal rudiment.
GnRH-immunoreactive perikarya were observed in the following regions: olfactory bulb and tubercle, vertical and horizontal limbs of the diagonal band of Broca, medial septum, medial preoptic and suprachiasmatic areas, anterior and lateral hypothalamus, and different regions of the hippocampus (indusium griseum, Ammon's horn). In addition to the known GnRH-pathways (preoptico-terminal, preoptico-infundibular, periventricular), we also observed GnRH-immunopositive processes in several major tracts and areas of the brain, including the medial and cortical amygdaloid complex, stria terminalis, stria medullaris thalami, fasciculus retroflexus, medial forebrain bundle, indusium griseum, stria longitudinalis medialis and lateralis, hippocampus, periaqueductal gray of the mesencephalon, and extracerebral regions, such as the lamina cribrosa, nervus terminalis and its associated ganglia.
The morphology and connections of the indusium griseum (IG) and anterior hippocampal continuation (AHC) suggest that this cortex contains analogues to several portions of the hippocampal formation.
At 22 days the handled animals showed a deficit in glial cells in both the anterior limb of the anterior commissure and in the indusium griseum. At 6 months of age the handled group contained more glial cells in both the rostral part of the anterior limb of the anterior commissure and the indusium griseum. It was noted that the timing of the handling coincided with the period of normal rapid proliferation of oligodendrocytes in the anterior commissure and of astrocytes in the indusium griseum..
Two indirect immunohistological methods (PAP and immunofluorescence) allowed them to visualize, for the first time, structures containing specific DSIP-like immunoreactivity in some areas of the rat brain: indusium griseum, nucleus septi lateralis, hippocampus, striae longitudinales of Lancisi , bandeletta diagnalis of Broca, pallidum, hypothalamus, hypophysis and neocortex. Some DSIP pathways seem likely: (1) indusium griseum - striae longitudinales - hippocampus; (2) nucleus septi lateralis - striae longitudinales , bandeletta diagonalis - hippocampus; (3) neurons of the pyramidal layer of the hippocampus - gyrus dentatus; (4) pallidum - commissura of Ganser - hypothalamus.
The authors detected LH--RH-containing neurons in the dorsal part of the hippocampus and in the indusium griseum of the rat.
These arise predominantly from cells in the LEA and project forward along the angular bundle to the piriform and periamygdaloid cortices, as well as the endopiriform nucleus, the lateral, basolateral, and cortical amygdaloid nuclei, the nucleus of the lateral olfactory tract, the olfactory tubercle, the anterior olfactory nucleus, the taenia tecta, and the indusium griseum.
The neostriatum, indusium griseum and anterior commissure of the mouse brain were examined by semithin light microscopy and electron microscopy in an attempt to identify and follow the development of the cells named neurolipomastocytes by Ibrahim et al. These cells were first seen at 5 days post natum in the indusium griseum and in the leptomeninges adjacent to it.
The glia:neuron ratio increased between 5 and 9 months in the neostriatum and indusium griseum and thereafter remained constant until 18 months-of-age. Between 18 and 22 months the glia:neuron ratio did not change in the neostriatum, but increased significantly in the indusium griseum due to a combination of a loss of neurons and an increase in glia. The increase in glia in the indusium griseum between 18 and 22 months was due to an increase in both astrocytes and microglia. Astrocytes first contained lipofuscin at 6 months in the neostriatum, at 9 months in the indusium griseum and at 15 months in the anterior commissure. Oligodendrocytes first contained lipofuscin at 12 months in the anterior commissure, at 18 months in the indusium griseum and at 18 months in the neostriatum. Foamy pericytes were found in all regions: from 6 months in the neostriatum; from 9 months in the indusium griseum and from 15 months in the anterior commissure.
Several new histological findings in six cases of the trisomy D1 syndrome are described: hyperplasia of fetal structures (indusium griseum, median raphe of the medulla oblongata) and completely developed cerebellar cortical heterotopia in the dentate nucleus.
A quantitative study of changes in total cell number was carried out in the indusium griseum and anterior commissure from fetal life to old age in the mouse brain. The changes in the number of mitotic and pyknotic cells were recorded in the indusium griseum, anterior commissure, subependymal and ependymal layers over the same period. The number of neurons which are produced and which migrate to the indusium griseum are in excess of the number eventually required and the surplus neurons are lost by cell death in late gestation and early postnatal life while synaptogenesis and neuronal differentiation is taking place. Mitotic activity in the indusium griseum levels out at 3 months postnatum with mitotic and pyknotic cells present in roughly equal numbers thereafter. Cell number in the indusium griseum and anterior commissure is fairly constant between 3 and 9 months, but glial number begins to decrease in all parts of the anterior commissure from 12 to 22 months. In the indusium griseum the number of glia increased slightly between 6 and 22 months.
The number of neurons and the number of glia in the indusium griseum did not change significantly. A similar increase in mitosis was found at 5 days postinjection in the indusium griseum. At 7 days postinjection a significant decrease occurred in pyknotic cells in the anterior commissure and indusium griseum.
An autoradiographic study of neuronal and glial production was carried out in the indusium griseum of mice.
The rostral part of the anterior limb of the anterior commissure (RAL) and the indusium griseum of the brains of Gunn rats and Wistar rats were examined quantitatively to assess the effects of hyperbilirubinaemia on the developing nervous system. The number of glia in the indusium griseum was similar in both groups but the Gunn rats showed a decrease of about 20% in the number of neutrons in this region.
Neurogenesis and gliogenesis in the indusium griseum were studied in semithin sections and with the electron microscope.
A quantitative study of the indusium griseum was carried out in control mice and in mice subjected to hypoxia at 390 mmHg for 2 days.
A quantitative study of the development of the indusium griseum was carried out in mice. It was not possible to separate the indusium griseum from the cingulate cortex until birth. The length of the indusium griseum increased from 1114+/-151 micrometer at birth to 2444+/-54micrometer at 25 days and 2544+/-80 micrometer at 140 days. Although the mean number of cells in equivalent sections decreased with age the distribution of cells was similar at all ages, with the majority of cells lying in the rostral half of the indusium griseum..
The indusium griseum was investigated quantitatively in 5-mo-old and 18-mo-old mice.
After injection of 3H corticosterone into adrenalectomized male rats, radioactivity was found to be selectively concentrated in neurons of septum, hippocampal complex (precommissural hippocampus, cornu Ammonis, gyrus dentatus, subiculum) indusium griseum, amygdala and in certain areas of the cortex.
Areas surrounding retrosplenial cortex in the posterior cingulate region were also evaluated in Nissl and Golgi preparations including the indusium griseum, subiculum (dorsal to the corpus callosum) and area 23. The transition from one cellular layer in the indusium griseum to five cellular layers in area 23 is made by the addition of layers II, III, IV and VI in retrosplenial cortex to the one ganglionic layer of the indusium griseum and subiculum. First, the structure of pyramidal cells progressively changes from those in the indusium griseum which have predominently round or oval somata and a preponderance of apical and few basal dendrites to those in layer V of retrosplenial cortex and area 23 which have pyramidal shaped somata and a great number of basal dendrites which branch frequently and spread horizontally for hundreds of microns. Evidence was not found that the indusium griseum, dorsal subiculum or layer V of retrosplenial granular cortex contain a significant number of stellate or fusiform cells.
After intraventricular injection, globus pallidus, indusium griseum, hippocampus, corpus callosum, and meninges.
One hour after the injection, radioactivity was found to be selectively concentrated in specific neurons of the septum, the hippocampal complex (precommissural hippocampus, cornu Ammonis, gyrus dentatus, subiculum), the indusium griseum, the amygdala and in certain areas of the cortex.
-
[ View All ]